 Para
entender mejor los conceptos y terminología relacionados con el
superenrollamiento, emplearemos como ejemplo una molécula bicatenaria
de DNA
Para
entender mejor los conceptos y terminología relacionados con el
superenrollamiento, emplearemos como ejemplo una molécula bicatenaria
de DNA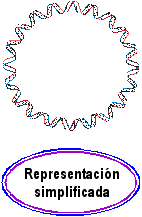 Ahora supongamos que flexionamos la molécula, aproximando sus extremos
para formar un círculo. Puesto que el número de vueltas de
hélice en la molécula es entero (10 en este ejemplo), los
extremos de ambas hebras quedarán correctamente alineados con posibilidad
de unirse por sendos enlaces fosfodiéster
Ahora supongamos que flexionamos la molécula, aproximando sus extremos
para formar un círculo. Puesto que el número de vueltas de
hélice en la molécula es entero (10 en este ejemplo), los
extremos de ambas hebras quedarán correctamente alineados con posibilidad
de unirse por sendos enlaces fosfodiésterAl ser esta molécula plana y sin tensión estructural (pues sigue adoptando la conformación B), se la denomina una forma relajada. Por definición, la conformación relajada es la que tiene superenrollamiento cero. (Estrictamente, lo que es plano no es la molécula, sino la trayectoria del eje de la doble hélice.)
Como consecuencia del cierre de
la molécula, las hebras![]() ya no se pueden separar físicamente (por ejemplo, tirando de ellas
en sentidos contrarios), sino que están trabadas, ligadas, entrelazadas
(similarmente a los eslabones de una cadena o los anillos de los prestidigitadores).
Concretamente, en este ejemplo están entrelazadas 10 veces, y se
dice que la molécula tiene un índice
de ligazón
ya no se pueden separar físicamente (por ejemplo, tirando de ellas
en sentidos contrarios), sino que están trabadas, ligadas, entrelazadas
(similarmente a los eslabones de una cadena o los anillos de los prestidigitadores).
Concretamente, en este ejemplo están entrelazadas 10 veces, y se
dice que la molécula tiene un índice
de ligazón![]() de 10 (L=10, o Lk=10,
del inglés linking number).
de 10 (L=10, o Lk=10,
del inglés linking number).

Un valor Lk = 0 significa que las dos hebras no están ligadas, es posible separarlas. Cuando Lk es distinto de cero las dos hebras están ligadas físicamente, es decir, no se pueden separar; esto es así a pesar de que no hay enlaces covalentes entre ellas; para referirnos a esta unión física sin enlace covalente se dice que hay un enlace topológico.
En una molécula con enlace topológico sólo se pueden separar las hebras mediante ruptura de un enlace covalente (fosfodiéster) o, dicho de otro modo, cortando una hebra. Lo mismo es aplicable para cualquier proceso que cambie el valor Lk (por ejemplo, de Lk=10 a Lk=9). Por la misma razón, la deformación, plegamiento, etc. de las hebras, sin cortarlas, no altera el índice de ligazón.
Si, en lugar de unir los extremos
de la molécula lineal tal y como quedaron enfrentados, antes rotamos
uno de ellos sobre sí mismo, de modo que se reduzca el enrollamiento
de la doble hélice en una vuelta (la torsión es menor, Tw=9),
y entonces unimos los extremos, tendremos una molécula con sus hebras
menos entrelazadas, con menor índice de ligazón, Lk=9. Al
mismo tiempo, los 104 pb ahora se reparten en sólo 9 vueltas de
hélice, por lo que en lugar de una vuelta cada 10,4 pb habrá
una cada 11,56 pb (=104/9) y la conformación no será ya de
tipo B. Esto supone una inestabilidad para la molécula, una tensión
estructural (similar a la que tiene el cable retorcido del teléfono
si intentamos aplanarlo sobre la mesa sin que forme bucles). La tensión
se puede liberar si la molécula forma un bucle![]() sobre sí misma, una vuelta superhelicoidal; así recupera
la torsión más estable (Tw=10, con 10,4 pb/vuelta). Al número
de vueltas superhelicoidales se le denomina retorcimiento
sobre sí misma, una vuelta superhelicoidal; así recupera
la torsión más estable (Tw=10, con 10,4 pb/vuelta). Al número
de vueltas superhelicoidales se le denomina retorcimiento![]() (W o Wr, del inglés writhe).
En este caso, debe considerarse de signo negativo (Wr=1), pues suple la
disminución de torsión inicial. El giro de esta superhélice
es dextrorso, hacia la derecha, como se puede comprobar con un modelo de
cuerda (la cuerda empleada como modelo tiene 4
hebras en lugar de 2, pero sirve para analizar el fenómeno):
(W o Wr, del inglés writhe).
En este caso, debe considerarse de signo negativo (Wr=1), pues suple la
disminución de torsión inicial. El giro de esta superhélice
es dextrorso, hacia la derecha, como se puede comprobar con un modelo de
cuerda (la cuerda empleada como modelo tiene 4
hebras en lugar de 2, pero sirve para analizar el fenómeno):
Forma relajadaModelo de DNA dextrorso (forma B) en conformación estable y sin
superenrollamiento
|
||
Alteración experimental hipotética:
Superenrollamiento negativo |
Alteración experimental hipotética:
Superenrollamiento positivo |
|
| Hélice infraenrollada, menos vueltas de
hélice de lo esperado;
Tw < Two aparece una tensión (no es la conformación estable). Más pb/vuelta que el modelo B-DNA (p.ej., 11). Menor giro por cada residuo (p.ej., 32°). |
Hélice sobreenrollada, más vueltas de hélice
de lo esperado;
Tw > Two aparece una tensión (no es la conformación estable). Menos pb/vuelta que el modelo B-DNA (p.ej., 9). Mayor giro por cada residuo (p.ej., 40°). |
|

Tw < Two , Wr = 0 (curva plana)
Ahora ya no hay tensión, Tw es de nuevo igual a Two;
este aumento que ha sufrido Tw ha tenido lugar gracias al cambio en el
retorcimiento, Wr ha disminuido:
|
Tw > Two , Wr = 0
Ahora ya no hay tensión, Tw es de nuevo igual a Two;
esta disminución que ha sufrido Tw ha tenido lugar gracias al cambio
en el retorcimiento, Wr ha aumentado:
|
|
| Puedes ver la generación de estos superenrollamientos en segmentos de vídeo: | ||
|
Superenrollamiento negativo
|
Superenrollamiento positivo
|
|
En un caso general, con reducción o aumento de varias vueltas de hélice, la tensión no tiene por qué eliminarse exclusivamente mediante retorcimiento (negativo o positivo, respectivamente), sino que en parte puede asumirse adoptando una conformación de menor o mayor torsión. Por ello, se cumple que cualquier cambio de Lk se puede repartir entre un cambio de torsión y un cambio de retorcimiento:
![]()
En el ejemplo anterior, al añadir inicialmente
el giro adicional antes de unir los extremos de la molécula, se
cambió de Lk=10 (conformación relajada) a Lk=9 (conformación
tensionada); la situación en ese momento era
Lk=9, Tw=9, Wr=0
y después del retorcimiento era
Lk=9, Tw=10, Wr=1
Dicho de otro modo, la eliminación de una ligazón o enlace
topológico, ![]() Lk=1,
se compensa:
Lk=1,
se compensa:
Puesto que ambas situaciones son interconvertibles, se puede considerar que hay superenrollamiento tanto cuando se forman superhélices (retorcimiento, curvatura del eje de la doble hélice) como cuando se mantiene una torsión diferente de la más estable (deformación de la geometría local de la doble hélice).
Puedes reforzar estas ideas visitando otra página que repite algunas cosas de forma más resumida, con comparaciones y con ejemplo numérico.
*Explicación:
![]() la conformación "B" del DNA posee 10,4 pb por cada vuelta de
doble hélice, con lo que 104 pb constituirán 104/10,4 = 10,0
vueltas.
la conformación "B" del DNA posee 10,4 pb por cada vuelta de
doble hélice, con lo que 104 pb constituirán 104/10,4 = 10,0
vueltas.
 |
 |
 |