

3 Superenrollamiento del DNA en la naturaleza
Algunas imágenes para ilustrar el superenrollamiento de las moléculas
circulares de DNA:
DNA circular superenrollado, sin ramificaciones
(la línea representa la doble hélice):

superhélice dextrorsa
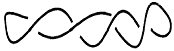
superhélice sinistrorsa
J.F.Marko (1997) Physica A 244, 263-277. |

DNA circular superenrollado, con una ramificación |
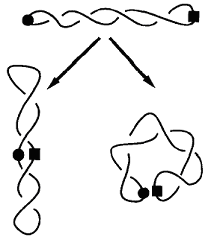
El superenrollamiento puede permitir que posiciones lejanas en la secuencia
se aproximen
|
|
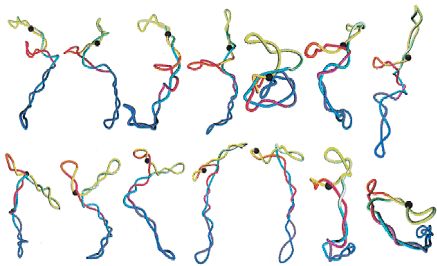
Modelos de DNA circular superenrollado
J.Huang, T.Schlick, A.Vologodskii (2001) Proc.Nat.Acad.Sci.USA
98, 968-973. |
 |
|
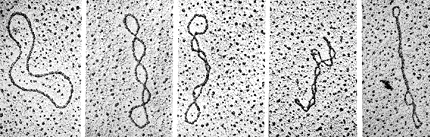
Observación al microscopio del superenrollamiento de un
DNA circular
D.Brutlag, Stanford Univ. [curso "Biochemistry
201" ] ] |
 |
¿Qué hace que las moléculas de DNA en las células
adopten conformaciones superenrolladas, y qué trascendencia biológica
tiene este fenómeno?
Hemos visto cómo el superenrollamiento
se origina cuando se giran las hebras en torno al eje de la doble hélice,
para perder o ganar torsión, y se unen de nuevo los extremos. En
las células, existen enzimas que catalizan precisamente este proceso.
Se las denomina topoisomerasas, pues
convierten unos topoisómeros[1]
en otros. Hay topoisomerasas de tipo I, que hidrolizan un enlace
fosfodiéster en una hebra, hacen pasar la otra hebra a través
del corte y vuelven a unir los extremos de la primera; como resultado,
el índice de ligazón de la molécula de DNA aumenta
o disminuye en una unidad. Otras topoisomerasas, denominadas de tipo
II, hidrolizan sendos enlaces fosfodiéster en ambas hebras y
hacen pasar otra doble hebra a través del corte, resellando éste
de nuevo; de este modo, el índice de ligazón varía
en dos unidades de una sola vez [2].
El resultado de la acción continua
de las topoisomerasas en las células es que el DNA se encuentra
habitualmente en estado superenrollado. Esto cumple varias funciones:
-
Las dimensiones de una molécula de DNA extendida exceden con mucho
las de la célula procariótica, el núcleo o el orgánulo
eucariótico que la deben contener. Mediante el superenrollamiento,
las moléculas de DNA se compactan y ocupan un volumen inferior en
varios órdenes de magnitud.
-
El superenrollamiento también permite que se acerquen regiones de
la molécula alejadas en su estructura primaria
 (estructura lineal del polímero), y esto es clave en procesos como
la regulación de la replicación
y la transcripción.
(estructura lineal del polímero), y esto es clave en procesos como
la regulación de la replicación
y la transcripción.
-
Del grado de compactación local conseguido depende la regulación
de la accesibilidad a la información genética y, por consiguiente,
la regulación de la expresión de los genes (transcripción).
-
Para que puedan tener lugar los procesos de replicación (copiado
del DNA) y transcripción (copiado de la información del DNA
en una molécula de RNA) es necesario que se separen las dos hebras
complementarias, lo cual supone un desenrollamiento local de la doble hélice.
Éste mediado por proteínas específicas, principalmente
polimerasas
y helicasas
provoca sobreenrollamiento en las regiones adyacentes. Además, dificulta
el desenrollamiento adicional, necesario para la progresión de los
citados procesos a lo largo de la molécula.
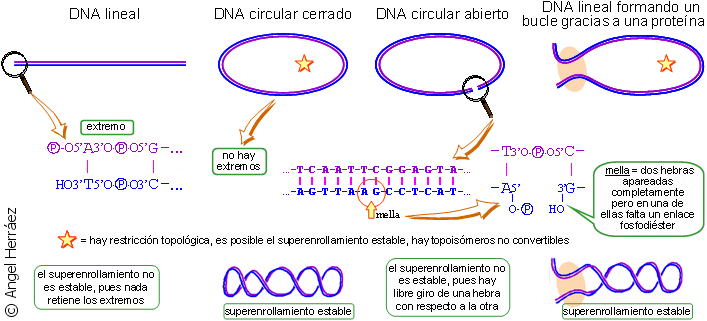
En la naturaleza, las moléculas circulares
de DNA (el cromosoma bacteriano, los plásmidos bacterianos y algunos
DNA víricos) se encuentran habitualmente superenrolladas. Como ya
se habrá comprendido por la exposición anterior, esa naturaleza
circular cerrada da lugar a los diferentes estados de superenrollamiento,
estables y no interconvertibles (salvo mediante rotura de enlaces covalentes).
En cuanto al material genético nuclear eucariótico, está
formado por grandes moléculas de DNA (cromosomas)
lineales, es decir, con dos extremos, que, por sí mismas, no pueden
mantener un superenrollamiento. Sin embargo, se encuentran asociadas estrechamente
con proteínas que, bien anclan los extremos delimitando un bucle
donde la doble hélice tiene el giro restringido, bien fuerzan el
enrollamiento de la molécula de DNA alrededor de la propia proteína,
formando uno o varios bucles superhelicoidales. En conclusión, tanto
en el DNA circular
como en el lineal
se da el fenómeno de superenrollamiento.
Algunas características adicionales de las formas superenrolladas
del DNA:
Superenrollamiento negativo del DNA
|
|
Superenrollamiento positivo del DNA
|
| Superhélice a derechas, dextrorsa. |
|
Superhélice a izquierdas, sinistrorsa. |
Se encuentra de forma natural en las eubacterias.
En eucariotas es más frecuente en las zonas codificantes(facilita
que se abra la doble hebra para la transcripción).
En eucariotas, transitoriamente por detrás de la RNApol durante
la transcripción (hélice infraenrollada, desenrollada). |
|
Se puede encontrar (aunque no es muy frecuente) en arqueobacterias.
En eucariotas, transitoriamente por delante de la RNApol
durante la transcripción (hélice sobreenrollada, apretada). |
| Une preferentemente intercalantes
que desenrollan la doble hélice (p.ej., bromuro de etidio y cloroquina). |
|
Une preferentemente ligandos que aprietan la doble hélice
(p.ej., netropsina). |
1. Topoisómero:
un isómero, típicamente de una molécula grande y compleja,
que es diferente de los otros debido a su topología; por ejemplo,
por el grado de retorcimiento o por el entrelazado de estructuras anulares.
Por ejemplo, los topoisómeros del DNA se diferencian en su índice
de ligazón Lk; un diferente valor de Lk da lugar a un distinto grado
de superenrollamiento de la doble hélice en cada topoisómero.
 (A Dictionary of Biology, Oxford University Press, © Market
House Books Ltd 2000 www.xrefer.com)
(A Dictionary of Biology, Oxford University Press, © Market
House Books Ltd 2000 www.xrefer.com)
2. Topoisomerasas: véase
Texto
Ilustrado de Biología Molecular e Ingeniería Genética,
Luque, p.152
o Bioquímica, 3ª ed,
Mathews et al., p.1017 Figura
24.30
Figura
24.31


